Paläo-Ökologie der Keuperflechten
Das Erdaltertum endet vor ca. 250 Millionen Jahren mit einem drastischen Einschnitt in die Lebewelt unseres Planeten. An der Wende Perm/Trias ist im fossilen Report ein weltweites Massenaussterben erkennbar, das einen Großteil der Arten auslöscht. Allein 96 % der marinen Arten verschwinden für immer (Raup 1991). Warmwasserfaunen, Plankton und Pilze trifft es am stärksten. Das Aussterben vollzieht sich nicht schlagartig und global, sondern in regionaler Differenzierung über Jahrmillionen hinweg und es ist schwierig, das Erlöschen von Arten mit sedimentologisch nachweisbaren geoökologischen Ereignissen und Veränderungen in einen ursächlichen Zusammenhang zu bringen. Solche Wendepunkte in der Lebewelt des Planeten und auch der unglücklich gewählte Begriff „Massenaussterben“ verführen leicht zur Vorstellung gewaltiger Katastrophen, die mit einem Schlag eine Epoche beenden und eine neue herbeiführen. Es wäre besser, von einem gegenüber dem Hintergrundsterben beschleunigten Aussterben zu sprechen. Sicher gab es mit dem Einschlag größerer Meteoriten oder großräumiger Vulkantätigkeit Szenarien in der Erdgeschichte, die Teile der Erdoberfläche verwüsteten. Aber die geologischen Spuren solcher dramatischen Ereignisse sind nicht unbedingt zeitlich korrelierbar mit tiefgreifenden Veränderungen im fossilen Report der Lebewelt (Bloos, 1999). Die Ursachen für das Massenaussterben von Arten sind eher komplex und stellen sich bei näherer Untersuchung als eine Art Kettenreaktion ganz unterschiedlicher Folgewirkungen dar, die größere Zeiträume beanspruchen. Dies gilt in besonderem Maße auch für die Perm-/Trias-Wende: Schwankungen des planetaren Klimas, Abkühlungen und Erwärmungen durch Änderungen in der Zusammensetzung der Atmosphäre, ausgelöst von weiträumiger vulkanischer Aktivität (vulkanischer Winter), Norddrift und Abkühlung der Pangäa (Kozur, 1999). Manche dieser Vorgänge stehen wohl auch in einem Zusammenhang mit dem allmählichen Zerbrechen des Superkontinentes.
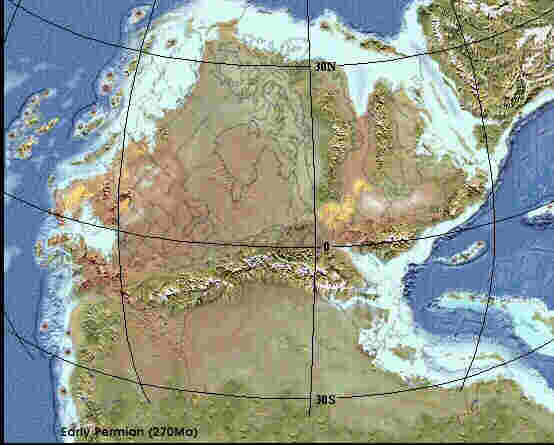
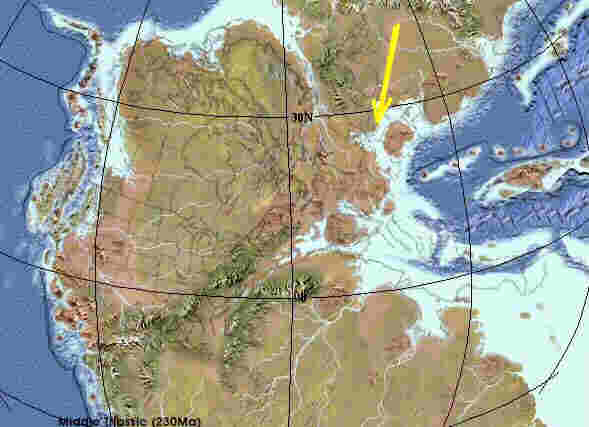
Nördliche Pangäa im Perm vor 270 Mio. Jahren nördliche Pangäa zur Keuperzeit vor 230 Mio. Jahren (Quelle 1)
Mit Beginn der Trias kommt es auf der Pangäa zu Rift-Vorgängen, die sich nachhaltig auch auf die Entwicklung des Germanischen Beckens auswirken. Im Perm (linke Karte) und der Unteren Trias (Buntsandstein) war das Gebiet festländisch, bedeckt mit kontinentalen Wüsten. Die Riftvorgänge, aus denen der Atlantik hervorgehen wird, führen auch im Kontinentalbereich zu Krustendehnungen und Absenkungen unter Meereshöhe. Folge ist die Überflutung großer Kontinentalflächen und Entstehung flacher Meeresbecken, in denen es unter ariden Klimabedingungen zur Ablagerung von Evaporiten kommt, das sind Eindampfungssedimente, wie Steinsalz, Gips und Anhydrit, die sich bei starker Evaporation (Verdunstung) aus übersättigten Lösungen absetzen. Eine kontinentale Senkungszone ist auch das Germanische Becken, der Sedimentationsraum, in dem der Keuper entsteht. Die rechte Karte zeigt das Germanische Becken (Pfeil) in der Mittleren Trias. Das Becken ist zum Teil vom Muschelkalkmeer überflutet. Im folgenden Keuper kommt es zu Regressionen und Transgressionen des flachen Randmeeres und Deltaschüttungen durch Ströme vom Baltischen Schild, Fennoskandien und dem Vindelizischen Massiv.
Massenaussterben als Evolutionsschub für die Flechtensymbiose
Das Aussterben von Arten durch großräumige Umweltänderung muss auch aus populationsgenetischer Sicht differenziert gesehen werden. Es bringt zwar vorübergehend eine Verminderung der Artenzahl, führt aber langfristig eher zu deren Steigerung . Man darf davon ausgehen, dass infolge des beschleunigten Aussterbens zu Beginn der Trias viele Lebensräume ausgeräumt oder nur schwach besiedelt sind. Das Angebot an differenziertem Lebensraum mit freien Ökonischen muss als starker Evolutionsschub wirken. Es kommt zu dem, was Evolutionsbiologen mit Flaschenhalseffekt, Gendrift und adaptiver Radiation beschreiben. So kann aus einer weiträumig , bis auf wenige, isolierte Restpopulationen vernichteten Art ein ganzer Schwarm neuer Spezies hervorgehen. Es gibt weniger Fressfeinde und Konkurrenten, die genetische Balance geht verloren, neue Formen haben mehr Chance auf Überleben als in einem während langer Zeiträume ausbalancierten Gleichgewicht. So können sich neue Tier- und Pflanzengruppen ausbreiten, wie etwa die frühen Dinosaurier, erste Säuger, Süßwassermuscheln oder die Bennettiteen (heute ausgestorbene Nacktsamer). Das gilt insbesondere auch für die Pilze, deren Massenentfaltung nach Kozur (1999) unmittelbar vor der Perm-/Trias-Wende endet. Pilze sind heterotroph, leben als Parasiten und Saprophyten. In einer Welt am Ende des Perm, deren Lebensräume nur schwach besiedelt sind, haben es auch die heterotrophen Pilze schwer. Begünstigt sind nun solche Pilz-Formen, die sich über das Zusammenleben mit Algen die Autotrophie erschließen. Es ist die einzigartige Vielfalt an kaum besetzten Ökonischen im Germanischen Becken zur Keuperzeit, in der die Evolution der Flechtensymbiose begann. Die Anatomie der fossilen Keuperflechten dokumentiert diesen Übergang. Der Bauplan rezenter Blatt-Flechten ist trotz morphologischer Vielfalt recht einheitlich. Die Keuperflechten sind dagegen morphologisch eher eintönig, lassen dafür aber verschiedene Baupläne erkennen, die es heute nicht mehr gibt (Eine Abhandlung hierüber ist in Vorbereitung.). Die Entstehung und parallele Entfaltung verschiedener Konstruktionen ist typisch für die Frühphase einer neuen Lebensform, wie es die Flechtensymbiose im Keuper ist. Entscheidend für die Einschätzung phylogenetischer Trends ist die Kenntnis der Umweltbedingungen, in denen sich eine solche Entwicklung vollzieht. Daher wird im Folgenden die groß- und kleinräumige Ökologie der Flechtenentstehung beschrieben, soweit das anhand der einschlägigen Literatur und eigenen Beobachtungen zur Zeit möglich ist.
Großräumige Ökologie des Germanischen Beckens
Orographie und Hydrologie: Der großräumige Charakter des Germanischen Beckens zur Keuperzeit lässt sich etwa so beschreiben: Das Germanische Becken ist eine allseitig von Hochländern und Gebirgen umrahmte Senkungszone (Fig. 1,2,3), die bereits über riesige Zeiträume hinweg als Sedimentationsraum gewaltige Mengen Erosionsmaterial aufgenommen hat. Seine Sedimentationsgeschichte im Keuper vollzieht sich als ein wechselvolles hydrologisches Geschehen, in dessen Verlauf sich das Becken abwechselnd mit Meer- oder Süßwasser füllt und immer wieder ganz oder teilweise austrocknet. Ursachen sind die Lage in Meereshöhe, niedrige Randschwellen der Meerespforten im Norden (zum Nordpolarmeer), im Westen (zum Protoatlantik), im Südwesten (Burgundische Pforte zur Tethis) und im Südosten (Böhmisch-Mährische Pforte zur Tethis). Hinzu kommen Meeresspiegelschwankungen und ein extremes Monsunklima. Die Festlandsgebiete Mitteleuropas liegen damals südlicher als heute, im Bereich der nördlichen Randtropen. Das Klima ist im Sommer subtropisch warm und wegen der Abschirmung durch Hochländer sowie der Entfernung zum Weltmeer kontinental geprägt, d.h., die Winter sind kalt und trocken. Durch ihre gewaltige Ausdehnung beiderseits des Äquators entwickelt die riesige Landmasse einen Megamonsun (Parrish, 1999) mit ausgeprägten Regen- und Trockenzeiten. Im Nordsommer liegt über dem aufgeheizten Germanischen Becken eine Tiefdruckzone, die feuchte Meeresluft aus dem Westen und Süden ansaugt und für reichlich Niederschläge sorgt. In Zeiten der Meeres-Regression (Meeresrückzug, Landwerdung), die im Keuper dominieren, bilden sich in den tiefergelegenen, teils abflusslosen Beckenregionen ausgedehnte Seenlandschaften (Fig. 1). Im Nordwinter dagegen kehrt sich die Windrichtung um. Die aus dem Becken nach Süden strömenden Luftmassen dörren das Land aus. Monatelang regnet es nicht, die Seen trocknen aus (Fig. 2). Wasser gelangt nur noch über Fremdlingsflüsse aus den umliegenden Gebirgen in das Becken, wo es in Salzseen verdunstet. Die monsunbedingte Hydrologie des Beckens wird überlagert von Meeresspiegelschwankungen der umgebenden Ozeane. In Zeiten der Meeres-Transgression, d.h. wenn das Meer in das Becken vordringt, entsteht ein seichtes Schelfmeer. Im Sommer wird es durch die hohen Niederschläge und Zuflüsse brackig, in der ariden Zeit dagegen verdunstet Meerwasser, werden Evaporite abgelagert. Daher kommt es in den Gewässern des Beckens zu häufigem Milieu-Wechsel zwischen süß, brackig und salinar, was sich nachhaltig auf die Lebewelt auswirkt. Das hydrologische Geschehen bedingt auch eine starke Dynamik in der Ausbildung des Reliefs. Von den Rändern her und teilweise auch aus weit entfernten Erosionsräumen, wie dem Baltischen Schild oder Fennoskandien, tragen mächtige Ströme eine Unmasse von Sedimenten in das flache Meeresbecken (Wurster, 1964). Damit gehören auch alluviale Schwemmfächer und Deltaschüttung zum Fazies-Repertoir des Keupers im Germanischen Becken.
1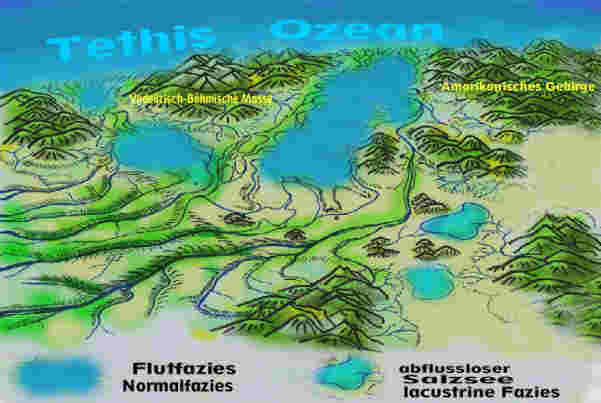 2
2 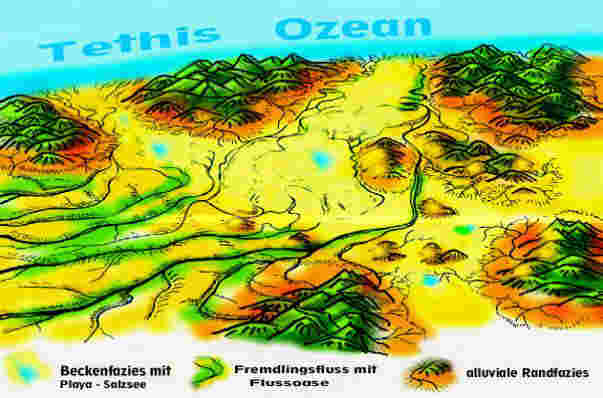
Klima und
Vegetation
Das Germanische Becken erhält durch das monsunale Regime der Pangäa ein extrem wechselfeuchtes Klima. Auch der jährliche Temperaturgang ist extrem. Im Sommer heiß durch die Nähe zu den Tropen, im Winter eher kühl durch die Beckenlage hinter abschirmenden Hochländern. Wasser erhält das Becken vor allem über Fremdlingsflüsse aus den umrahmenden Hochländern und Gebirgen, in denen die starken Monsunregen als Steigungsregen niedergehen. Die Feuchte wird modifiziert durch das Relief. Regionen, die nicht geflutet werden, erhalten nur Niederschlagswasser, das aber durch die hohe Verdunstung nur kurze Vegetationszeiten gestattet. Die längste Zeit im Jahr sind diese Gebiete wüstenartig. Im Winter dehnen sich die Wüsten sogar noch aus, weil das Becken über die abschwellenden Fremdlingsflüsse weniger Wasser erhält und geflutete Teile austrocknen. So grenzen im Germanischen Becken Lebensräume aneinander, die sich in ihrer Ökologie stark unterscheiden: Immerfeuchte Flussoasen und sumpfige Niederungen, die ausreichend Wasser speichern und in den Dürrezeiten nicht austrocknen, wechselfeuchte Regionen, die sich jahreszeitlich begrünen und aride Räume mit wüstenhaftem Charakter, in die Oasen eingestreut sind. Die Vielfalt der Lebensräume und Biotope ermöglicht einerseits die Besiedelung mit zahlreichen Arten, stellt aber andererseits durch ihre starke Dynamik die Lebewelt vor besondere Probleme. Das ökologische Repertoire reicht von der kontinental geprägten lebensfeindlichen Salzwüste bis hin zu immerfeuchten sumpfigen Niederungen mit subtropischem Charakter. Die kargen ariden Festländer sind eine „moderne Welt“ der Reptilien und Koniferen, die ihre mesozoische Erfolgsstory mit der Eroberung arider Lebensräume beginnen. Die subtropischen feuchten Niederungen gehören noch den archaischen Schachtelhalmen, Farnen und Amphibien, wie Mastodonsaurus, die ihre Blütezeit bereits hinter sich haben. Alles in allem eine Ökologie und Lebewelt, wie sie gegensätzlicher nicht sein kann. Die Reste von Farnen, Schachtelhalmen, Cycadeen, Nadelhölzern und zahlreichen anderen meist ausgestorbenen Arten sind in den Taphozönosen (Fossilgesellschaften) des Keupers häufig vertreten und wecken den Wunsch, daraus das Bild einer Flora oder Vegetation zu entwerfen. Kelber (2001) nennt „vier markante Florenkomplexe“ für den gesamten Keuper, wobei man sich unter dem Begriff „Florenkomplex“ die Gesamtheit der Taphofloren verschiedener Fundstellen vorzustellen hat: Die Floren des Werksandsteins, des Schilfsandsteins, des Coburger Sandsteins und des Rhät.
Fossile
Keuper-Floren?
In der paläobotanischen Literatur ist es üblich, die Aufsammlungen fossiler Pflanzen einer oder mehrerer zeitlich korrelierbarer Fundstellen als fossile Floren oder Taphofloren zu bezeichnen. Die Anwendung des Begriffs „Flora“ auf die Vergesellschaftung fossiler Pflanzenreste, wie sie im Begriff „Taphoflora“ zum Ausdruck kommt, ist allerdings problematisch, weil sie zu falschen Vorstellungen führen kann. Bei Anwendung auf die heutige Pflanzenwelt versteht man unter Flora den gesamten Artenbestand eines bestimmten Gebietes zu einer gegebenen Zeit. Sie wird mit einer Artenliste erfasst. Sofern die Lebensansprüche der enthaltenen Arten bekannt sind, ermöglicht die Flora auch begrenzte Rückschlüsse auf die Ökologie ihres Lebensraumes. Eine Flora ist stets eingebunden in eine Vegetation. Mit dem Vegetationsbegriff werden Mengenanteile und Artenkombination, d.h. die Vergesellschaftung der Arten erfasst. Die floristische Zusammensetzung einer Vegetation hängt ab von Ökofaktoren, wie Klima, Boden oder Relief, und von der ökologischen Valenz der Arten. Sie ist deshalb ein wichtiger Indikator für die Umweltbedingungen, in der sich die Vegetation entfaltet. Ihre Aussagekraft hängt ab von der Verlässlichkeit der Artenliste und der Häufigkeit der einzelnen Arten. Bei rezenten Pflanzenbeständen sind Artenpräsenz (floristische Daten), Vergesellschaftung und Mengenanteile (vegetationskundliche Daten) vor Ort relativ leicht erfassbar. Dagegen führt der Versuch, eine nicht mehr existierende Vegetation anhand ihrer fossilen Reste zu rekonstruieren, in große Unsicherheiten, sowohl hinsichtlich der Artenliste als auch der Mengenanteile.
Flora bedeutet die zeitgleiche Präsenz bestimmter Pflanzenarten in einem definierten Gebiet. Ob „Taphofloren“ diese Kriterien erfüllen, erscheint fraglich, vor allem was die Zeitgleichheit betrifft. Die Artenliste einer fossilen Fundstelle ist ein floristischer Ausschnitt einer Vegetation unbekannter räumlicher Ausdehnung. Auch Vegetationsdaten sind auf Fossilbasis unsicher, weil Verwesungsvorgänge, zufällige Verteilung und Einbettung der Pflanzenreste die ursprünglichen Häufigkeiten und Mengenverhältnisse der Arten verändern können. Aufsammlungen verschiedener Fundstellen liefern daher wohl mehr einen Floren-Querschnitt für eine Raum-Zeit unbestimmbarer Größe. Ein Florenquerschnitt ist aber ein Durchschnittswert, dem weder eine konkrete Flora noch eine konkrete Umwelt zugeordnet werden können. Für eine paläo-ökologische Analyse sind pflanzliche Makroreste daher kaum geeignet.
Betrachten wir als Beispiel den Keuper. Er umfasst insgesamt ca. 24 Millionen Jahre Sedimentationsgeschichte des Germanischen Beckens. Mit der Gliederung dieses Zeitraumes befasst sich die Stratigrafie, die hierbei verschiedene Verfahren zur Anwendung bringt. Eine wichtige Methode ist die Lithostratigrafie, die anhand des Gesteinsaufbaus nach Merkmalen, wie Korngröße, Sedimentfärbung, Geochemie, Schichtgefüge und Fossilgehalt gliedert. Doch es wird sehr schwierig, wenn es darum geht, eine bestimmte Schicht weiträumig zu verfolgen. „Abgesehen von Einzelnachweisen biostratigrafisch verwertbarer Fossilien im Unteren und Mittleren Keuper, kann nur im Oberen Keuper biostratigrafisch anhand von Ostrakoden, Lamellibranchiaten und Megasporen gegliedert werden“ (Bachmann et al. 1999). Diese für die Fauna ernüchternde Beurteilung gilt in noch stärkerem Maße für die Möglichkeiten einer stratigrafischen Gliederung anhand pflanzlicher Makroreste. Die Vegetationsdecke ist zu keiner Zeit des Keupers geschlossen und über größere Distanzen einheitlich. Wir müssen auf Grund der ökologischen Verhältnisse davon ausgehen, dass sie ein Mosaik von Pflanzengesellschaften mit sehr unterschiedlicher floristischer Zusammensetzung ist, das sich zeitlich und räumlich relativ schnell wandelt. Von dieser Vegetationsentwicklung, die immerhin unvorstellbare 24 Millionen Jahre umfasst, erfahren wir gegenwärtig über fossile Reste aus einigen Dutzend Fundstellen, die räumlich über die Keuperformation verteilt und zeitlich schwer korrelierbar sind. Gemessen am Zeitraum und dem wechselvollen ökologischen Geschehen erscheint jeder Versuch, Florenbilder des Keupers zu entwerfen, mit großen Unsicherheiten behaftet und weitgehend spekulativ. Selbst die Aufsammlungen einer Fundstelle geben nur Anhaltspunkte für ein konkretes Florenbild zu einer bestimmten Zeit im Keuper, denn heute nicht mehr erkennbare Sedimentations- und Erosionsereignisse mit unterschiedlichen Geschwindigkeiten können sich seit der Fossilwerdung ereignet haben. Wenige Zentimeter Sediment zwischen zwei fossilen Blättern können bedeuten, dass diese Pflanzen Zeitgenossen waren und rasch nacheinander eingeschlossen wurden. Es können aber auch Jahrtausende dazwischen liegen, wenn zwischen den Einbettungen die Sedimentation aufhörte oder sogar Erosion einsetzte. Beutler et al. (1999) bezeichnen den Keuper als einen „Stapel von Schichtlücken“ und vermuten, dass „nur 5-10 % der im Keuper enthaltenen Zeit durch Sedimente dokumentiert ist“. Welch dramatische Änderungen im Vegetationsbild bereits innerhalb weniger Jahrtausende oder nur Jahrhunderte ablaufen können, lehrt uns die gut untersuchte nacheiszeitliche Entwicklung der Vegetation in Europa.
Somit stellt sich grundsätzlich die Frage, ob überhaupt auf taphonomischem Wege ein Florenbild für eine bestimmte Formation gewonnen werden kann. Für makroskopische PflanzenFossilien muss das aus den genannten Gründen bezweifelt werden. Größere Aussicht, zu echten floristischen Daten zu kommen, besteht im Mikrobereich. Zu den hier beschriebenen Kutikeln und Pilzen kommen die im fossilen Pflanzenmoder häufigen Palynomorphen, wie Sporen und Pollen, als Leitfossilien und Zeitindikatoren hinzu. Trotz Bedenken wegen der unbekannten Größe des Einzugsgebietes, kann man mit einiger Sicherheit davon ausgehen, dass die Arten einer zusammengeschwemmten und damit allochthonen Palynomorphengemeinschaft, - das ist eine Ansammlung von Sporen, Pollen, Kutikeln u.a. Mikrofossilien, die nicht vom Fundort kommen, sondern durch Wind oder Wasser antransportiert werden - wenn auch räumlich getrennt, so zumindest zeitgleich gelebt haben. Mit dem Einsatz des Mikroskops bei der Fossiliensuche öffnet sich daher im fossilen Pflanzenmoder ein Fenster in die Lebewelt des Keupers, das nicht nur eine große Zahl bislang unbekannter Arten, sondern auch floristische Daten liefert. Hierzu Heunisch (1999): „Darüber hinaus bietet die unterschiedliche Zusammensetzung der Mikroflorengemeinschaften in den verschiedenen Ablagerungsbereichen die Möglichkeit, Schichten gleichen Alters faziesübergreifend miteinander zu vergleichen. Pflanzliche Mikrofossilien (Palynomorphe) liefern die Grundlagen für eine biostratigrafische Einstufung und faziesübergreifende Aussagen“.
Der Schilfsandstein (Stuttgartformation) ist mit geschätzten einer Million Jahre eine relativ kurze Zeitspanne im Sedimentationsgeschehen des Keupers. Umso mehr erstaunen uns die gewaltigen Ablagerungsmengen, die in dieser Epoche entstehen. Fluviatile Prozesse prägen den Schilfsandstein. Aus Schüttungsrichtung und Mineralgehalt der Sande ermittelte Wurster (1964) den Baltischen Schild und Fennoskandien als Liefergebiete des Erosionsmaterials und Ursprung der ins Germanische Becken vordringenden
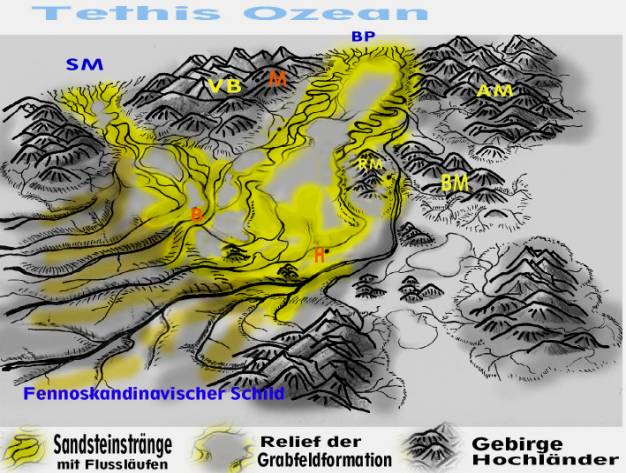 Fig. 3: Germanisches Becken zur Schilfsandsteinzeit mit Blickrichtung
Nord-Süd
Fig. 3: Germanisches Becken zur Schilfsandsteinzeit mit Blickrichtung
Nord-Süd
SM = Schlesisch-Mährische Pforte, BP = Burgundische Pforte, RM=Rheinisches Massiv, AM=Amorikanisches Massiv, VB=Vindelizisch-Böhmisches Festland, BM=Brabanter Massiv (zur Orientierung: H=Hamburg, B=Berlin, M=München).
 /
Fig. 4: mäandrierender und sedimentierender Fluss in
Alaska
/
Fig. 4: mäandrierender und sedimentierender Fluss in
Alaska
Flüsse. Man nimmt heute an, dass das Stromsystem des Schilfsandsteins anfangs den Tälern eines bereits in der Grabfeldformation angelegten Reliefs folgt, sich teilweise bis zu vierzig Meter tief in den liegenden Gipskeuper eingräbt und schließlich das gesamte Relief der älteren Grabfeldformation mit mächtigen Sedimentdecken verhüllt (Beutler et al. 1999). Damit ergibt sich für die Genese des Schilfsandsteins das Bild einer weiten, von niedrigen Landrücken durchzogenen Flusslandschaft( Fig. 1). Zahlreiche Ströme aller Größenordnungen durchqueren auf ihrem Weg zum Tethismeer mäandrierend und anastomoisierend das Germanische Becken, wobei sie ausgedehnte Sandstränge aufschütten, auf denen sie träge pendeln . Zum Vergleich, der auf seinem Sedimentationsbett pendelnde Citin-River in Alaska (Fig. 2). Das Monsunklima der Pangäa sorgt im Zusammenspiel mit dem Relief und den Fremdlingsflüssen aus weit entfernten Liefergebieten für eine komplexe Hydrologie im Germanischen Becken. Dabei kommt es zwischen den beiden Hauptjahreszeiten, der Regen- und der Trockenzeit, zu einem extremen Wandel der Lebensbedingungen: Im warm-feuchten Sommermonsun schwellen die Flussläufe mächtig an, ertränken große Teile des Beckens unter Schichtfluten und verwandeln es in eine riesige Seenlandschaft (Fig. 1). Die Wassermassen, die ins Becken gelangen, sind gewaltig, denn die Flüsse schleppen ihre Erosionsfracht quer durch das gesamte Becken bis zur Tethis. Auch die Salzseen, die nicht vom Stromsystem erreicht werden, erhalten Frischwasser aus den Hochländern am Beckenrand und verwandeln sich an ihren Ufern in Oasen. Es ist anzunehmen, dass sich, vergleichbar zu den heutigen Halbwüsten, in den nicht überfluteten Beckenregionen durch Niederschlagswasser kurzzeitig eine üppige Vegetation aus vornehmlich krautigen Pflanzen ausbreitet und Herden pflanzenfressender Saurier anlockt, wie z.B. frühe Dinosaurier aus der Gruppe der Plateosaurier. Im kühl-trockenen Wind des Wintermonsuns verdorrt das Land und das Becken wird zunehmend lebensfeindlich. Die Fremdlingsflüsse schwellen ab, kehren in ihre Rinnen zurück oder versiegen ganz. Seen trocknen aus, trockenrissige Tonebenen und Salzkrusten entstehen ( Fig. 2). Dazwischen persistieren in die Trockenzeit hinein immerfeuchte Flussoasen und sumpfige Niederungen, die ausreichend Wasser speichern und auch in den Dürrezeiten nicht austrocknen. Sie können für wassersuchende Saurier zur tödlichen Falle werden. Berühmtheit erlangten die „Plateosaurier-Friedhöfe“ von Trossingen, Halberstadt und Frick. Als Erklärung für die teils in Sterbestellung erhaltenen Skelette wird neben anderen Hypothesen ein gewaltsamer Tod durch Steckenbleiben im zähen Schlamm diskutiert (Sander, 1999). Die Tiere müssen auf ihren weiten Wanderungen zwischen den jahreszeitlich wechselnden Weidegebieten Trockenregionen durchqueren und geraten bei der Suche nach Wasser in Schlammlöcher oder Sümpfe, aus denen sie sich nicht mehr befreien können und verenden. Die extremen Jahresgänge von Niederschlag und Temperatur erzeugen im Zusammenspiel mit den Besonderheiten des Reliefs eine starke Dynamik und Variabilität der Standorte und damit der Ökonischen. So entsteht ein komplexes Selektionsmuster für hohe Anpassungsfähigkeit in den verschiedensten Tier- und Pflanzengruppen. Typisch sind artenarme aber individuenreiche Lebensgemeinschaften, wie z.B. in der Wirbellosen-Fauna. Viele Arten zeigen „eine beachtliche innerartliche Variationsbreite, was durch instabile Lebensbedingungen hervorgerufen wird ...“ (Seegis 1999) In der Tierwelt kann man eine „Becken-“ von einer „Randfauna“ unterscheiden. Trotz der wechselhaften Ökobedingungen können Tiere durch kurzfristiges Ausweichen in Feuchträume die extremen Trockenperioden überstehen und sich von dorther neu ausbreiten. Überhaupt müssen Wanderungen oder besondere Überlebenstechniken im Öko-Szenarium des Schilfsandsteins eine große Rolle gespielt haben. Vielleicht gehört zu ihnen auch die Entwicklung eines allseitig geschlossenen Pilzlagers. Die darin eingeschlossenen Algen ermöglichen als Glucoselieferanten dem Pilz die Emanzipation vom feuchteabhängigen Saprophyten zur autotrophen Flechte, die in aride Räume vorstößt.
Wechsel von Überflutung und Austrocknung, Erosion und Sedimentation durch Flussbettverlagerungen sorgen aber auch dafür, dass bereits zur Keuperzeit ein Großteil der Ablagerungen und damit der fossilen Zeugnisse wieder umgelagert, abtransportiert und völlig zerstört wird. Davon übrig bleibt ein zeitlich, wie räumlich äußerst lückenhaftes Mosaik lokaler Lagerstätten: Fossile Sümpfe, Fluss- und Seeufer, Flussrinnen, fossile Böden mit Spurenfossilen und zahlreichen Relikten tierischen und pflanzlichen Lebens. Es gibt kaum eine andere erdgeschichtliche Formation, die in der Vielfalt der Ablagerungen dem Schilfsandstein vergleichbar wäre. Die Rekonstruktion konkreter Biozönosen wird durch die räumliche und zeitliche Diskontinuität der fossilen Fundstätten stark erschwert, wenn nicht sogar unmöglich. Hinzu kommen im ganzen Becken verbreitete, deutlich erkennbare Zyklizitäten in der Sedimentation. Wenn etwa ein See austrocknet, kommt es zu einer charakteristischen vertikalen Schichtung. Die so entstehende Schichtfolge, die lacustrine Fazies, wiederholt sich mit jeder Abfolge von Seebildung und Verlandung. Da über das Germanische Becken räumlich verteilt zahlreiche Seen gleichzeitig verdunsten und sich das ganze über Jahrmillionen hin wiederholt, entsteht eine gewaltige Anzahl lokaler Zyklen. Auch im Falle des fossilen Moders kann man davon ausgehen, dass der Transport im Wasser zur Entstehung der Fundstelle beiträgt. Vielleicht ist es ein Altwasser, ein Durchbruchsfächer, irgendein Gleithang mit träger Strömung oder der Grund eines seichten Sees, auf dem die feuchte Pflanzenmasse zu Liegen kommt . Während das Wasser in der Sonnenhitze verdunstet, vermehren sich noch für kurze Zeit saprophytische Bakterien und Pilze in der Modermasse, bis alles Leben in der Trockenheit erstarrt. Sandflug oder das Sediment einer Schichtflut bedeckt die Stelle und entzieht sie dem weiteren biologischen Abbau. Die Taphozönose kann entstehen. Durch Erosion werden aber bereits zur Keuperzeit ca. 90 % der so gebildeten Sedimente wieder ausgeräumt und das nicht gleichmäßig, sondern in räumlich-zeitlicher Diversität. Die heute davon noch erhaltenen Sedimentationszyklen und ihre Fossilien zeitlich zu korrelieren, ist daher sehr schwierig. Angesichts dieser enormen Dynamik wird deutlich, dass es sich bei jeder Fundstelle im Schilfsandstein um eine Singularität handelt, die inselartig in Raum und Zeit isoliert, ein jeweils winziges Fenster öffnet in eine ausgestorbene Lebewelt.